VIVO Pathophysiology
Ontogeny of Small Intestinal Function
Survival depends on the ability of neonates to absorb nutrients beginning immediately after birth, and the intestinal mucosa is certainly functional at that time. However, the neonate's mucosa is 'fine-tuned' to exploit it's standard diet - milk - and changes in distinctive ways over time to become nearly adult-like at the time of weaning. Mice, humans and pigs have been the most studied with regard to ontogeny of mucosal function. Changes have been characterized for both digestive and absorptive processes.
Carbohydrate Digestion and Absorption
A well studied example of ontogenic change is in small intestinal disaccharidase activity. Lactose is the major, almost sole dietary carbohydrate for neonates, but, except for some humans, is not consumed by adult animals. On the other hand, neonates do not normally ingest sucrose or starch, but adults often do.
Neonates and adults alike are unable to absorb dietary disaccharides and require brush border disaccharidases to hydrolyze those molecules into absorbable monosaccharides. Considering these facts, it would make physiologic sense for neonatal intestine to have high lactase activity, but low sucrase and maltase activities, and for lactase to diminish dramatically and sucrase/maltase to increase by the time of weaning.
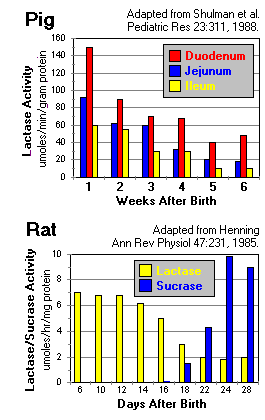
Indeed, this is exactly what happens in rodents, pigs and, for the most part, humans. In both pigs and humans, lactase activity is first observed early in gestation (e.g. 8-9 weeks in human gestation) and rises rather dramatically during the third trimester to achieve high levels during the first week of life.
The top figure to the right depicts brush border lactase concentrations in different regions of the neonatal pig intestine, from one through six weeks of age. Note the steady decline in lactase concentration throughout the small intestine, a pattern seen in essentially all animals studied.
The bottom figure to the right shows the inverse relationship between lactase and sucrase concentrations in the small intestine of young rats. In humans, the change in sucrase expression is less dramatic, and sucrase concentrations are relatively stable from birth through adulthood.
There are numerous differences in dissacharidase activity among species. As another example, and in contrast to rats, cattle do not express small intestinal sucrase activity at any time during development.
The mechanisms responsible for these developmental changes in intestinal disaccharidase expression remain poorly defined. Changes in diet clearly play some role, but do not appear to be the major regulator. For example, prolonging the period of suckling in rats causes a delay in decline of lactase expression, but does not alter the timing of increased sucrase expression. A postnatal rise in secretion of glucocorticoids also seems to modulate expression of disaccharidase genes, but again, is clearly not the major controller. Treating young rats with the glucocorticoid corticosterone slightly accelerated the rise in sucrase expression, but did not affect lactase expression.
Hexoses are absorbed by the same transport mechanisms in neonates and adults and there are not major quantitative changes in capacity for transport. For example, in both neonates and adults glucose is transported from the lumen via the sodium-dependent hexose transporter. Expression of this protein does not undergo significant changes during postnatal development in many animals, but in ruminants, there is a dramatic decrease in expression of this transporter (SGLUT1) during the first few weeks after birth, and very low levels of expression after weaning. As might be expected from changes in sucrase expression, fructose transport is low during the suckling period and rises significantly at weaning.
Protein Digestion and Absorption
Mechanisms of protein digestion and absorption are distinctly different between neonates and adults. Brush border peptidases appear to be present in the neonatal mucosa, but their expression does increase substantially at weaning. Also, amino acid transporters are functional at birth, although there may be quantitative changes as the animal matures.
Most dramatically however, the neonatal intestine efficiently transports intact proteins across the mucosa for varying periods of time after birth. In rodents, there is minimal secretion of gastric and pancreatic proteases in the neonatal period. Mucosal epithelial cells pinocytose intact proteins and degrade them in lysosomes. One important exception to this pathway is seen in absorption of immunglobulins, which bind to Fc receptors and are transcytosed across the epithelium instead of being targeted to lysosomes. Neonatal pigs seem to use a slightly different strategy - their proteolytic systems are more intact, but intestinal contents are rich in protease inhibitors (presumably from serum), which also allow intact proteins to reach the epithelium for pinocytosis. Finally human neonates also absorb macromolecules intact and this ability persists well into childhood, but the mechanisms allowing it is not well studied.
Other Developmental Changes
In neonatal rodents, calcium is absorbed by a nonsaturable, vitamin D-independent process. By the end of weaning, calcium transport is adult-like, in that it saturable, dependent on vitamin D and concentrated in the duodenum.
References and Reviews
- Freeman TC, Wood IS, Sirinathsinghji DJ, etc: The expression of the Na+/glucose cotransporter (SGLT1) gene in lamb small intestine during postnatal development. Biochimica et Biophysica Acta. 146:203, 1993.
- Henning SJ: Ontogeny of enzymes in the small intestine. Ann Rev Physiol 47:231-245, 1985.
- Henning SJ, Rubin RC, Shulman RJ: Ontogeny of the intestinal mucosa. In: Physiology of the Gastrointestinal Tract, LR Johnson (ed), Raven Press, pp 571-610, 1994.
- Le Huerou I, Guilloteau P, Wicker C, etc. Activity distribution of seven digestive enzymes along small intestine in calves during development and weaning. Dig Dis Sci. 1992; 37:40-46.
- Shulman RJ, Henning SJ, Nichols BL: The miniature pig as an animal model for the study of small intestinal enzyme development. Pediatric Res 23:311-315, 1988.
Send comments to Richard.Bowen@colostate.edu